Rôle de la myéline
La myéline est une membrane biologique graisseuse constituée
d’une épaisse couche de lipides et de protéines qui entoure les axones des
neurones. Elle permet de protéger les axones contre l'agression des (exo)nucléases, c'est-à-dire d'enzymes, et accélère la circulation des
potentiels d'action qui s'apparentent aux messages nerveux. Les oligodendrocytes sont les cellules du système nerveux
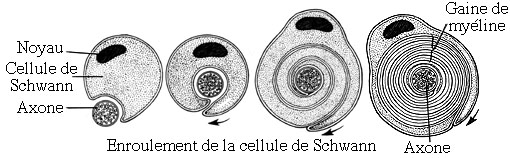
central qui permettent l’élaboration de la gaine de myéline. En effet, la
gaine de myéline résulte de l’enroulement des prolongements cytoplasmiques des
oligodendrocytes autour des axones. Cette gaine peut atteindre une épaisseur
allant jusqu’à 40 feuillets superposés.


Elles correspondent à des zones de faible résistance électrique riches en canaux-sodium et canaux-potassium et représentent donc les zones privilégiées pour le déclenchement des potentiels d'action (conduction de l’influx nerveux). La conduction nerveuse le long de l'axone myélinisé s'effectue de façon passive, comme si l’influx nerveux « sautait » d'un nœud de Ranvier à l'autre. Le rôle des nœuds de Ranvier est donc de créer et de transférer le potentiel d’action le long des neurones tandis que le rôle de la myéline est d’accélérer la circulation du message nerveux.
Gaines de myéline (microscope électronique à balayage)

{kind=link}
Schéma montrant l'enroulement de l'oligodendrocyte autour d'un axone
{kind=link}
La myéline forme une gaine conductrice très efficace : les lipides, hydrophobes, empêchent aux éléments aqueux d’interagir avec les axones et donc de perturber les influx électriques qui circulent dans le SNC. Il existe naturellement des parties axonales non recouvertes de myéline. Ces parties se nomment les nœuds de Ranvier.
Elles correspondent à des zones de faible résistance électrique riches en canaux-sodium et canaux-potassium et représentent donc les zones privilégiées pour le déclenchement des potentiels d'action (conduction de l’influx nerveux). La conduction nerveuse le long de l'axone myélinisé s'effectue de façon passive, comme si l’influx nerveux « sautait » d'un nœud de Ranvier à l'autre. Le rôle des nœuds de Ranvier est donc de créer et de transférer le potentiel d’action le long des neurones tandis que le rôle de la myéline est d’accélérer la circulation du message nerveux.
Conséquences de la
démyélinisation
L’absence de la gaine de myéline entraîne un ralentissement
considérable de la propagation des messages nerveux et la disparition des nœuds
de Ranvier.
Dans le cas de la sclérose en plaques, les cellules
immunitaires s’attaquent à la myéline et la dégrade jusqu’à sa destruction.
Ainsi, des pans entiers d’axones se retrouvent dénudés ce qui perturbe voire
empêche la circulation de l’influx nerveux. Les zones les plus touchées par la
démyélinisation se situent généralement dans l’encéphale et le nerf optique.
L'inflammation s'installe, active des cellules immunitaires
qui se comportent comme en cas de plaie; cela génère alors une « croûte » ou
une « plaque ».

Dégradation de la gaine de myéline
{kind=link}
Au regard de la diversité des formes évolutives de la
maladie, des analyses ont été effectuées sur quelques échantillons de patients
pour connaître les conséquences métaboliques d’une démyélinisation.
Lésions métaboliques
La SRM (Spectrométrie par Résonance Magnétique) permet de
détecter les lésions métaboliques qui caractérisent la sclérose en
plaque :
- Une inflammation : augmentation du lactate dans le milieu extracellulaire
- Une démyélinisation : augmentation de la choline dans le milieu extracellulaire
La choline est l’une des principales molécules qui composent la phosphatidylcholine. Celle-ci se trouve en quantité importante dans les membranes cellulaires des neurones. Ainsi, l’augmentation du taux de choline dans le cerveau traduit une altération des cellules neuronales qui peut être due notamment à la dégradation de la myéline puisque la perte de phosphatidylcholine est détectable par la libération de choline dans le milieu extracellulaire.
- Disfonctionnement axonal : diminution de N-Acétyl-Aspartate
N-acétyl-aspartate est synthétisé à partir d’aspartate et de
coenzyme acétyl A dans les neurones. NAA est ensuite transporté des neurones
vers le cytoplasme des oligodendrocytes où les enzymes aspartoacylase clivent
le groupement acétate pour produire des acides gras et stéroïdes de synthèse.
Ceux-ci sont ensuite utilisés pour former la myéline qui entoure les neurones. La
diminution de N-acétyl-aspartate dans le CNS traduit donc une démyélinisation.
Antigène(s) de la
myéline
Plusieurs études ont été menées pour tenter d’identifier
l’antigène de la gaine de myéline visé par les lymphocytes T. L’une de ces
études, concentrée sur l’identification des récepteurs présents sur les
lymphocytes T (TCR) complémentaires de leur cible ont obtenu des résultats
intéressants. En effet, les analyses du sang de patients atteints de SEP ont
révélé la présence de récepteur TCR dont la séquence génétique est complémentaire à
celle de la protéine basique de la myéline appelée MBP (Oksenberg JR, Panzara
MA, Begovich AB, Mitchell D, Erlich HA et Murray RS, Selection for T-cell receptor Vb-Db-Jb gene rearrangements with
specificity for a myelin basic protein peptide in brain lesions of multiple
sclerosis, Nature 1993).
Plus récemment, d’autres études scientifiques ont trouvé des
preuves montrant l’attaque d’autres antigènes de la myéline, de cellules
gliales ou de neurones, notamment MOG (Myelin Oligodendrocyte Glycoprotein) et PLP (Protéine Proteo Lipidique), deux autres antigènes.
Ainsi, il n’est pas encore possible de définir clairement un
unique antigène visé par les lymphocytes auto-réactifs de la sclérose en plaques.
Aucun commentaire:
Enregistrer un commentaire